Our favorite
morphogen system is the dorsal-ventral (DV) axis specification in the Drosophila melanogaster (fruit fly) embryo. In this system, a protein called "Dorsal" has a high concentration (in the nucleus) on the ventral side of the embryo and a low concentration on the dorsal side. (By the way, if you're wondering why it seems the name is backwards -- why is Dorsal absent on the dorsal side, but present on the ventral side? -- you might want to read our post
here.) It also directs the cells along the DV axis to express different genes, in a concentration dependent fashion. So Dorsal acts as a morphogen.
If you read our post introducing the idea of a
morphogen, you know that the typical way a morphogen concentration gradient forms is by a small set of cells producing the protein, then diffusion causes the concentration of that protein to spread out. This is not the case with Dorsal. It is present everywhere, but cannot act because it's bound to an inhibitor protein called Cactus. (Yes, seriously, it's called Cactus. Again, you might want to read about
why fruit fly gene names are so weird.)
But, on the ventral side of the embryo, a signal from the receptor Toll causes Cactus to be degraded, which releases Dorsal, allowing it to go into the nucleus and direct gene expression (see the above figure). This "Dorsal nuclear concentration gradient" (see figure below) is one of the earliest signals to direct the cells in the fly embryo to differentiate into different types, specifying (from ventral to dorsal) muscle, neuron, skin, and an extraembryoinc tissue called the amnioserosa. So the Dorsal gradient is very important, and is one of the more well-studied morphogen systems.
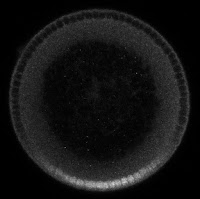 |
| The Dorsal nuclear concentration gradient: This is a cross section of a 2.5 hr old fruit fly embryo that has been treated so that Dorsal protein fluoresces. Dorsal protein is concentrated in the embryo's nuclei on the ventral side (bottom), and depleted on the dorsal side (top). |
